Sung-Gyu Lee,
Hyun Kang ![]()
For correspondence:- Hyun Kang Email: hkang@dankook.ac.kr Tel:+82415501452
Received: 18 November 2015 Accepted: 20 February 2016 Published: 31 March 2016
Citation: Lee S, Kang H. Anti-neuroinflammatory effects of ethanol extract of Inula helenium L (Compositae). Trop J Pharm Res 2016; 15(3):521-526 doi: 10.4314/tjpr.v15i3.13
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the in vitro antioxidant and anti-neuroinflammatory activities of Inula helenium extract (IHE) against lipopolysaccharide (LPS)-induced production of nitric oxide (NO) by primary microglial cells.
Methods: Cell viability was estimated by 3-(4, 5-dimethylthiazol-2-yl)-2, 5- diphenyl-tetrazolium bromide (MTT) assay. Antioxidant activity was evaluated using 1, 1-diphenyl-2-picryl-hydrazyl (DPPH) radical scavenging assay. LPS-stimulated BV-2 microglia were used to study the ex
Results: Pretreatment with IHE prior to LPS treatment significantly inhibited excessive production of NO (p < 0.001 at 20, 40, 80 and 100 μg/mL) in a dose-dependent manner, and was associated with down-regulation of ex
Conclusion: IHE may be beneficial in preventing and treating neurodegenerative and oxidative stress-related diseases.
Introduction
Inula helenium, also known as elecampane, is commonly found in China. In traditional medicine, it is extensively used primarily for treatment of abdominal pain, emesis, diarrhea, and threatened abortion [1]. In addition, the roots are effective in the treatment of diuretic, diaphoretic, expectorant, and anthelmintic remedy [2]. I. helenium has been identified that I. helenium contains sesquiterpene lactones and flavonoids [3–7], while other studies have reported that phenolic compounds exist in the Compositae [8]. However, till date, the effect of I. helenium extract on microglia-mediated neuroinflammatory diseases has not been reported.
Microglia are the resident macrophage-like cells of the central nervous system (CNS) with a broad role in the brains innate immunity and in inflammatory neuro-pathologies [9]. They are highly responsive to stress and injury and become immediately and focally activated in response to Alzheimer’s disease (AD), Parkinson’s disease (PD), multiple sclerosis (MS), and amyotrophic lateral sclerosis (ALS) [10,11]. Microglia are involved in initiating inflammatory responses in the brain through secreting a variety of inflammatory mediators, including nitric oxide (NO), inducible NO synthase (iNOS), interleukins (IL) and tumor necrosis factor (TNF)-α [11]. Thus, modulation of activated microglia is an effective strategy for therapeutics in the neuro-inflammation.
The most common cause of inflammatory responses is an exposure to lipopolysaccharide (LPS), a component of the outer membrane of Gram-negative bacteria [12]. LPS-activated BV-2 microglia cells responses induction and secretion of pro-inflammatory mediators. Therefore, suppression of aberrant activation of BV-2 microglia cells may have valuable therapeutic potential for the treatment of inflammatory diseases such as AD and PD [13].
In this study, we investigated the effects of I. helenium on the NO production, expression of iNOS, and pro-inflammatory cytokines from LPS-stimulated BV-2 microglia cells. The findings show that I. helenium may be used for the prevention or treatment of cerebral inflammatory diseases.
Methods
Preparation of Inula helenium extract (IHE)
Inula helenium collected at the end of June to about the middle of August were obtained from the local market, Seoul, South Korea. Inula helenium was washed in running tap water, dried at 60 °C for 24 h and then ground to fine powder. Each powder was extracted with 10 volumes (v/w) of 70 % ethanol at a room temperature for 72 h and filtrated. The filtered extract was concentrated by a rotary evaporator (EYELA NVC-2000, Tokyo, Japan) under reduced pressure and lyophilized (Jeol Ltd, Tokyo, Japan).
DPPH radical scavenging activity
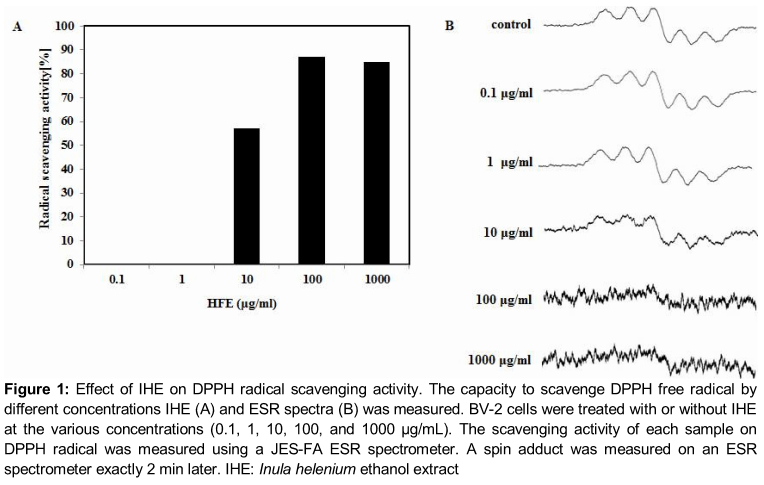
This assay was based on the scavenging of stable 2, 2-diphenyl-1-picrylhydrazyl (DPPH, Sigma-Aldrich, St Louis, MO, USA) radicals by the radical scavenging antioxidants in IHE. The radical scavenging capacity was evaluated by employing a reaction mixture constituted by aliquots of the IHE and a DPPH methanolic solution as described previously [14]. Briefly, a sample solution of 60 μl of each IHE, was added to 60 μl of DPPH (60 μM) in methanol. After mixing vigorously for 10 s, the mixture was transferred into a 100 µL Teflon capillary tube and the scavenging activity of each sample on DPPH radical was measured using a JES-FA ESR spectrometer (Jeol Ltd., Tokyo, Japan). A spin adduct was measured on an ESR spectrometer exactly after 2 min. Experimental conditions were as follows: central field, 3,475 G; modulation frequency, 100 kHz; modulation amplitude, 2 G; microwave power, 5 mW; gain, 6.3 x 105, and temperature, 298 °K.
Cell cultures and viability
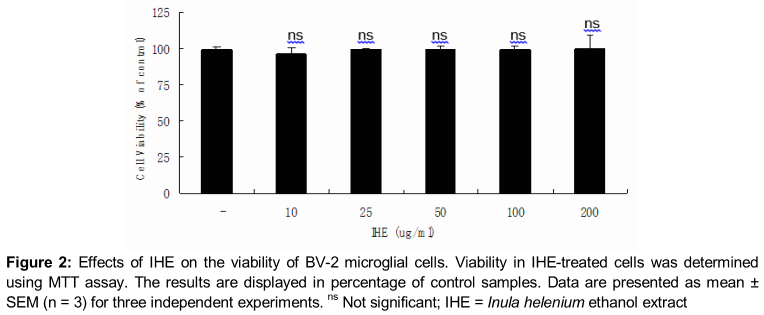
BV-2 microglia cells were grown in RPMI1640 (Invitrogen, Carlsbad, CA, USA), supplemented with 10 % FBS (Hyclone, Logan, UT, USA) containing 100 U/ml of penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen) in 37 °C in humidified atmosphere of 5 % CO2. In all experiments, cells were pre-treated with the indicated concentrations of IHE for 1 h before the addition of LPS (1 µg/ml, Sigma-Aldrich, St. Louis, MO, USA) in serum free RPMI1640. An equal volume of IHErile water was added to all control treatments. Cell viability was determined by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay as described previously [15]. Cells were incubated with various concentrations of IHE for 24 h followed by MTT for 4 h, and then 100 μL of isopropanol (in 0.04 N-hydrochloric acid) was added to dissolve the formazan crystals. The absorbance was read at 570 nm using the Anthos 2010 spectrophotometer (Salzburg, Austria). Cell viability was calculated as relative absorbance compared to control.
Nitric oxide assay
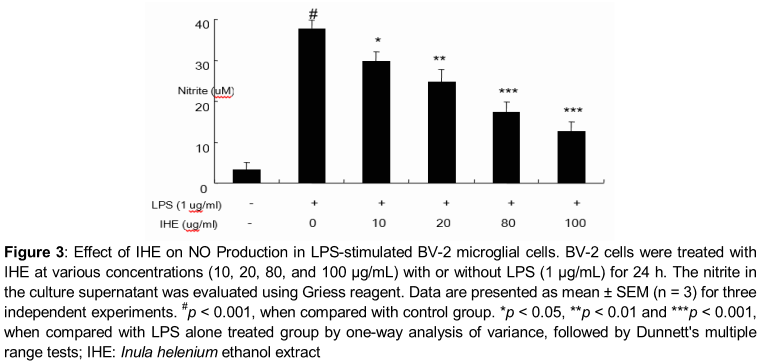
The amount of NO production in the medium was detected with the Griess reaction [16]. Each supernatant was mixed with the same volume of Griess reagent (1 % sulfanilamide in 5 % phosphoric acid and 0.1 % naphthyl ethylenediamine dihydrochloride in water). The absorbance of the mixture at 540 nm was determined with an ELISA reader (Bio-Tek Instrument, Winooski, VT, USA), nitrite concentration was determined using sodium nitrite as a standard.
Immunoblot analysis
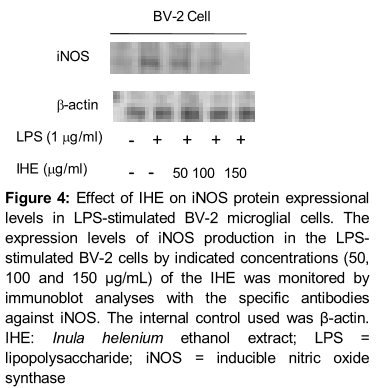
BV-2 cells were lysed in a buffer containing 20 mM Tris‐HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 50 mM β‐glycerophosphate, 1 % NP‐40, 1 mM Na3VO4 and 1 X protease inhibitor cocktail (Complete MiniTM, Roche, Mannheim, Germany). The lysates were centrifuged at 15,000 x g for 15 min and supernatants were collected. The concentrations of protein samples were measured using the BCA reagent (Pierce, Rockford, IL, USA). The protein was separated by sodium dodecylsulphate (SDS) polyacrylamide gel electrophoresis (PAGE) and transferred to PVDF membranes. After blocking with 5 % non‐fat milk, the membranes were hybridized with 1:1000 dilution of primary antibodies for iNOS and β-actin for 12 h at 4 °C. Membranes were washed with Tris‐Buffered Saline containing 0.1 % Tween‐20 (TBST) three times with 5 min interval and hybridized with horseradish peroxidase‐conjugated mouse or rabbit secondary antibodies for 1 h at room temperature. After washing three times with TBST, antigen‐antibody complexes were visualized with an ECL Western Blot Detection Kit (GE Healthcare) according to the manufacturer’s instructions. The antibodies against iNOS and β-actin were purchased from Cell Signaling Technology INC. (Beverly, MA, USA).
IL-6 assay
BV-2 microglia cells (1 x 105 cells/well) were cultured on 96 well plates and treated with the IHE at indicated concentrations for 1 h and stimulated with LPS (1 µg/ml). At 24 h post-LPS treatment, the production of IL-6 was determined in cell supernatant using assay kits (BD Biosciences, San Jose, CA, USA) as per the manufacturer’s instructions. The IL-6 assay was performed at 20 °C and optical absorbance was measured at 450 nm using ELISA reader.
Statistical analysis
All data are presented as the mean ± SEM (n = 3). Statistical analyses were performed using SAS statistical software (SAS Institute, Cray, NC, USA) using one-way analysis of variance, followed by Dunnett's multiple range tests. P < 0.05 was considered statistically significant.
Results
Effect of IHE on DPPH radical scavenging activity
As shown in A, IHE exhibited significant DPPH radical scavenging activity in a dose-dependent manner showing a maximum effect at 1 mg/mL of concentration. The ESR spectroscopy data at 0.1, 1, 10, 100 and 1000 µg/mL is represented in B.
Effect of IHE on LPS-induced NO production
IHE treatment did not exhibit any significant cytotoxicity in BV-2 microglial cells treated for 24 h at concentrations up to 200 µg/ml, and in all cases the viability was found above 96 % by MTT assay (). As shown in , treatment of LPS resulted in the excessive production of NO. Pretreatment of IHE significantly suppressed the LPS-induced NO production in a concentration-dependent manner (). The maximum effect was shown at 100 µg/mL (p < 0.001).
Effect of IHE on LPS-induced expressional levels of iNOS
Western blot analysis showed that the protein expression of iNOS in the LPS-stimulated BV-2 cells was also suppressed in a concentration dependent manner. However, treatment with IHE exhibited a broad spectrum of inhibitory effect on the expression of iNOS induced by LPS in BV-2 cells ().
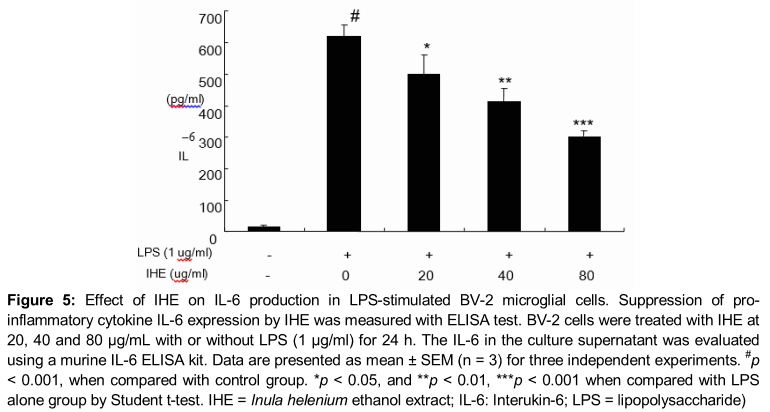
Effect of IHE on IL-6 production in LPS-stimulated BV-2 cells
As shown in , IL-6 levels increased significantly after LPS treatment (1 µg/ml) when compared to those in untreated cells (p < 0.001). However, IHE significantly inhibited IL-6 production in a concentration-dependent manner in LPS-stimulated BV-2 cells (p < 0.05 at 20 µg/mL and p < 0.01 at 40 and 80 µg/mL, respectively).
Discussion
The present study demonstrated that IHE possesses an anti-neuroinflammatory activity in LPS-stimulated BV-2 microglia cells and antioxidant activity. IHE also exhibited significant antioxidant activity. Free radicals and reactive oxygen species (ROS) are important causative factors in the development of age-related neuroinflammatory and neurodegenerative diseases [14]. Thus neutralization by antioxidants and radical scavengers can reduce neuroinflammation. It is well known that DPPH radical assay as one of the widely used methods for evaluating the free radical scavenging activities of several antioxidants in a relatively short period of time [16]. In our present study, IHE also exhibited significant free radical scavenging effect indicating that it might contain potential antioxidant agents. In our present study, the IHE exhibited significant free radical scavenging effect indicating that the Inula helenium might contain potential antioxidant agents.
In BV-2 microglia cells, NO is generated by the inducible isoform of NO synthase (iNOS) and iNOS are responsible for the symptoms of many neuro-inflammatory diseases [16]. Thus, high levels of NO are produced from L-arginine by iNOS in the brain by prolonged activation of microglial cells, and this response is associated with the progression of various neurodegenerative diseases [17]. Our results clearly showed that IHE attenuated LPS-induced iNOS expression and downstream NO production. This result indicates that IHE acts principally by regulating NO generation at the post-transcriptional level and could be beneficial for preventing the progression of neuro-inflammation by BV-2 microglial activation. Pro-inflammatory cytokines such as TNF-α are the initiators of the inflammatory response and the mediators of the development of chronic inflammatory diseases [18]. Therefore, the over-production of pro-inflammatory cytokines by activated microglia cell is a possible etiological factor of neurological disorders [19]. Microglial cell activation by LPS produces various cytokines including IL-6 leading to the attraction of neutrophils and the accumulation of neutrophil-secreted proteases and ROS at the site of inflammation. From our results, it appears that IHE may act by suppressing the expression of IL-6 protein levels and thereby inhibiting NO production and iNOS expression levels in LPS-stimulated BV-2 cells.
Conclusion
These data demonstrate that IHE has anti-inflammatory properties in LPS-induced BV-2 microglial activation through the down-regulation of inflammation-related gene expression, including iNOS and pro-inflammatory cytokine such as IL-6. Therefore, IHE can be considered as a useful therapeutic and preventive approach for the treatment of several neurodegenerative and oxidative stress-related diseases.
Declarations
Acknowledgement
References
Archives


News Updates